
Waarom hebben bacteriën flagella nodig? Flagella en hun locatie in bacteriën
Voor beweging in het aquatische milieu gebruiken sommige micro-organismen een flagellaatorgaan - "flagellum". Dit orgaan, ingebouwd in het celmembraan, zorgt ervoor dat het micro-organisme met een bepaalde snelheid naar believen in de gekozen richting kan bewegen.
Mannelijke voortplantingscellen gebruiken het flagellum ook voor beweging.
Gedurende een bepaalde tijd wisten wetenschappers van flagella. De kennis van hun structurele kenmerken, die pas in het laatste decennium of zo naar voren kwam, kwam voor hen echter als een enorme verrassing. Het bleek dat het flagellum door een zeer complexe "organische motor" beweegt en niet door een eenvoudig trilmechanisme, zoals eerder werd gedacht.
Deze motor is gevormd volgens dezelfde mechanische principes als een elektromotor. Het heeft twee hoofdonderdelen: een bewegend deel ("rotor") en een stationair deel ("stator").
Het bacteriële flagellum verschilt van alle organische systemen die mechanische bewegingen ondergaan. De cel gebruikt niet de beschikbare energie die is opgeslagen in de ATP-moleculen. In plaats daarvan heeft het een speciale energiebron: het micro-organisme gebruikt de energie van de stroom van ionen door hun buitenmembranen. De interne structuur van de motor is uiterst complex. Ongeveer 240 verschillende eiwitten zijn betrokken bij de aanmaak van het flagellum. Elk van hen neemt een bepaalde plaats in. Wetenschappers hebben ontdekt dat deze eiwitten signalen geleiden om de motor aan en uit te zetten, verbindingen vormen die beweging op atomair niveau vergemakkelijken en andere eiwitten activeren die het flagellum aan het celmembraan hechten. De modellen die zijn ontwikkeld om de werking van het systeem samen te vatten, zijn voldoende om de complexe structuur van het systeem te beschrijven. (1)

De complexe structuur van het bacteriële flagellum is op zichzelf al voldoende om de evolutietheorie te weerleggen, aangezien het flagellum een onherleidbare complexe structuur heeft. Zelfs als één enkel molecuul van deze ongelooflijk complexe structuur verdwijnt of beschadigd raakt, zal het flagellum niet werken en het micro-organisme niet ten goede komen. De flagellum zou vanaf het allereerste moment van zijn bestaan perfect moeten werken. Dit feit onderstreept nogmaals de absurditeit van de bewering van de evolutietheorie over "stapsgewijze ontwikkeling".

Zelfs die wezens die evolutionisten als 'de eenvoudigste' beschouwen, hebben een verbazingwekkende structuur. Het bacteriële flagellum is een van de talloze voorbeelden. Dit micro-organisme beweegt in water en beweegt dit orgaan op zijn schaal. Toen het interne systeem van dit bekende orgaan werd bestudeerd, waren wetenschappers over de hele wereld verrast toen ze ontdekten dat het micro-organisme een uiterst complexe elektromotor heeft. Deze elektromotor, die ongeveer vijftig verschillende moleculaire subeenheden omvat, heeft een nogal ingewikkelde structuur, zoals hieronder weergegeven.
Het bacteriële flagellum is een duidelijk bewijs dat zelfs zogenaamd "primitieve" wezens een ongewone structuur hebben. Naarmate de mensheid de details steeds beter begrijpt, wordt het duidelijk dat de organismen waarvan wetenschappers van de 19e eeuw, waaronder Darwin, dachten dat ze de eenvoudigste waren, in feite net zo complex zijn als andere. Met andere woorden, met de komst van een begrip van de perfectie van de schepping, wordt het duidelijk dat het geen zin heeft om te proberen een alternatieve verklaring voor de schepping te vinden.

Het micro-organisme drijft in een stroperig vloeibaar medium, roterende spiraalpropellers genaamd flagella.
Het bacteriële flagellum is een nanomachine die bestaat uit 25 verschillende eiwitten, in hoeveelheden van enkele tot tienduizenden. Het bestaat uit een verzameling van dit grote aantal eiwitten, die elk in verschillende delen een specifieke functie hebben, zoals motorrotatie, isolatie, aandrijfas, regelaarschakelvolgorde, universele koppeling, spiraalpropeller en een roterende versterker voor zelf- samenkomst.
Flagellaire eiwitten worden gesynthetiseerd in het cellichaam en getransporteerd langs een lang, smal centraal kanaal in het flagellum naar het perifere (externe) uiteinde, waar ze efficiënt en onafhankelijk complexe nanostructuren kunnen creëren met behulp van een flagellaire punt als aandrijfmotor. De roterende motor, waarvan de diameter slechts 30 tot 40 nm is, roteert het flagellum met een frequentie van ongeveer 300 Hz en een vermogen van 10-16 W, met een energieconversie-efficiëntie van bijna 100%.
De structurele ontwerpen en functionele mechanismen die worden gevonden in het complexe mechanisme van het bacteriële flagellum, kunnen de mensheid veel innovatieve technologieën bieden die de basis zullen vormen voor toekomstige nanotechnologie, waarvoor we veel nuttige toepassingen kunnen vinden.
De ontwikkeling van de microbiologie heeft de afgelopen decennia veel ontdekkingen opgeleverd. En een daarvan zijn de eigenaardigheden van de beweging van flagellaire bacteriën. Het apparaat van de motoren van deze oude organismen bleek zeer complex te zijn en, volgens het principe van zijn werking, heel anders dan de flagella van onze naaste eukaryote verwanten van de protozoa. De heetste controverse tussen creationisten en evolutionisten is losgebarsten rond de motor van flagellaire bacteriën. Dit artikel gaat over bacteriën, hun flagellaire motoren en nog veel meer.
Algemene biologie
Laten we om te beginnen onthouden wat voor soort organismen ze zijn en welke plaats ze innemen in het systeem van de organische wereld op onze planeet. Het Bacteria-domein verenigt een groot aantal eencellige prokaryotische (zonder gevormde kern) organismen.
Deze levende cellen verschenen bijna 4 miljard jaar geleden in de arena van het leven en waren de eerste kolonisten op de planeet. Ze kunnen heel verschillende vormen hebben (kokken, staafjes, vibrio's, spirocheten), maar de meeste zijn gevlagd.
Waar leven bacteriën? Overal. Er zijn er meer dan 5 × 10 30 op de planeet. Er zijn ongeveer 40 miljoen van hen in 1 gram aarde, tot 39 biljoen leven in ons lichaam. Ze zijn te vinden op de bodem van de Marianentrog, in hete "zwarte rokers" op de bodem van de oceanen, in het ijs van Antarctica, en op dit moment zijn er tot 10 miljoen bacteriën op je handen.
De waarde is onmiskenbaar
Ondanks hun microscopische grootte (0,5-5 micron), is hun totale biomassa op aarde groter dan de biomassa van dieren en planten samen. Hun rol in de circulatie van stoffen is onvervangbaar en hun eigenschappen van consumenten (vernietigers van organisch materiaal) laten niet toe dat de planeet wordt bedekt met bergen lijken.
Welnu, vergeet ziekteverwekkers niet: bacteriën zijn ook de veroorzakers van pest, pokken, syfilis, tuberculose en vele andere infectieziekten.
Bacteriën hebben toepassing gevonden in menselijke economische activiteiten. Van de voedingsindustrie (zuivelproducten, kazen, ingemaakte groenten, alcoholische dranken), de "groene" economie (biobrandstoffen en biogas) tot methoden van celengineering en de productie van medicijnen (vaccins, serums, hormonen, vitamines).

Algemene morfologie
Zoals eerder vermeld, hebben deze eencellige vertegenwoordigers van het leven geen kern, hun erfelijk materiaal (DNA-moleculen in de vorm van een ring) bevinden zich in een bepaald gebied van het cytoplasma (nucleoïde). Hun cel heeft een plasmamembraan en een dichte capsule gevormd door peptidoglycan-mureïne. Van de celorganellen hebben bacteriën mitochondriën, chloroplasten en andere structuren met verschillende functies.
De meeste bacteriën zijn flagellaat. Door een dichte capsule op het oppervlak van de cel kunnen ze niet bewegen door de cel zelf te veranderen, zoals amoeben doen. Hun flagellen zijn dichte eiwitformaties van verschillende lengtes en een diameter van ongeveer 20 nm. Sommige bacteriën hebben een enkele flagellum (monotrichs), terwijl andere er twee hebben (amphitrichs). Soms zijn de flagellen in bundels gerangschikt (lophotrichs) of bedekken ze het hele oppervlak van de cel (peritrichs).
Velen van hen leven als afzonderlijke cellen, maar sommige vormen clusters (paren, kettingen, filamenten, hyfen).

Bewegingsfuncties
Flagellaatbacteriën kunnen zich op verschillende manieren verplaatsen. Sommige gaan alleen vooruit en de richting wordt veranderd door salto's. Sommige kunnen trillen, terwijl andere bewegen door te schuiven.
Bacteriële flagella vervullen niet alleen de functies van een cellulaire "peddel", maar kunnen ook een "instaphulpmiddel" zijn.
Tot voor kort werd aangenomen dat de bacteriële flagellum kwispelt als de staart van een slang. Recente studies hebben aangetoond dat het bacteriële flagellum veel complexer is. Het werkt als een turbine. Bevestigd aan de aandrijving, draait deze in één richting. De aandrijving, of flagellaire motor van een bacterie, is een complexe moleculaire structuur die werkt als een spier. Met dit verschil dat de spier na de compressie moet ontspannen, en de bacteriële motor constant draait.

Nanomechanisme van het flagellum
Zonder diep in te gaan op de biochemie van beweging, merken we op dat tot 240 eiwitten betrokken zijn bij het creëren van de flagellaire aandrijving, die zijn onderverdeeld in 50 moleculaire componenten met een specifieke functie in het systeem.
In dit aandrijfsysteem hebben bacteriën een rotor die beweegt en een stator die deze beweging aandrijft. Heeft een aandrijfas, naaf, koppeling, remmen en gaspedaal
Met deze miniatuurmotor kunnen bacteriën in slechts 1 seconde 35 keer hun eigen grootte reizen. Tegelijkertijd besteedt het lichaam voor het werk van het flagellum zelf, dat 60 duizend omwentelingen per minuut maakt, slechts 0,1% van alle energie die de cel verbruikt.
Het is ook verrassend dat de bacteriën alle onderdelen van hun locomotiefmechanisme "on the fly" kunnen vervangen en repareren. Stel je voor dat je in een vliegtuig zit. En technici wisselen de bladen van een draaiende motor.
Flagellaatbacteriën versus Darwin
Een motor die kan werken met snelheden tot 60.000 tpm, zelfreinigend en alleen koolhydraten (suikers) als brandstof gebruikt, met een apparaat dat lijkt op een elektromotor - zou zo'n apparaat tijdens het evolutieproces kunnen zijn ontstaan?
Dit is de vraag die Michael Behe, doctor in de biologie, zichzelf in 1988 stelde. Hij introduceerde in de biologie het concept van een onherleidbaar systeem - een systeem waarin alle onderdelen tegelijkertijd nodig zijn om zijn werk te garanderen, en het verwijderen van ten minste één onderdeel leidt tot een volledige verstoring van het functioneren ervan.
Vanuit het standpunt van Darwins evolutie vinden alle structurele veranderingen in het lichaam geleidelijk plaats en worden alleen succesvolle door natuurlijke selectie geselecteerd.
De conclusies van M. Behe, vermeld in het boek "Darwin's Black Box" (1996): de motor van de flagellaire bacterie is een ondeelbaar systeem van meer dan 40 delen, en de afwezigheid van ten minste één zal leiden tot het volledig niet- functionaliteit van het systeem, wat betekent dat dit systeem niet door natuurlijke selectie had kunnen ontstaan ...

Balsem voor creationisten
De scheppingstheorie, zoals gepresenteerd door de wetenschapper en professor in de biologie, decaan van de Faculteit Biologische Wetenschappen aan de Lehigh University of Bethlehem (VS), M. Behe trok onmiddellijk de aandacht van kerkministers en aanhangers van de theorie van de goddelijke oorsprong van het leven.
In 2005 was er zelfs een proces in de Verenigde Staten waar Behe getuige was van de kant van de aanhangers van de theorie van "intelligent design", die de introductie van de studie van creationisme in Dover-scholen in de cursus overwoog " Over panda's en mensen." Het proces ging verloren, het onderwijzen van een dergelijk onderwerp bleek in strijd met de huidige grondwet.
Maar de controverse tussen creationisten en evolutionisten duurt vandaag voort.
Bacteriën zijn een vorm van organismen die op aarde zijn verschenen bij de oorsprong van de planeet.
Lange tijd waren bacteriën niet alleen de eerste, maar ook de enige bewoners van de aarde. Het lichaam van de bacil is eencellig, heeft geen exacte kern en is allemaal vaag, de bacteriën behoren tot de eenvoudigste eencellige, na verloop van tijd begonnen veel andere stoffen de aarde te bewonen, maar de structuur van de microbe blijft nog steeds de meest elementaire .
Volgens hun structuur zijn ze: staafvormig, bolvormig, ingewikkeld en spiraalvormig. Microben met een tweecellige lichaamsstructuur zijn zeldzaam.
De reproductie van micro-organismen vindt extreem snel plaats, in 3 uur kunnen meer dan 100 duizend bacteriën worden gevormd.
Voortplanting vindt plaats door de cel in 2 delen te verdelen, dan raakt het lichaam direct overgroeid met een schil en zo is het nieuwe lichaam klaar. Voor de voortplanting gebruiken microben een gunstige omgeving, maar hun eisen daarvoor zijn niet zo groot. Om de kweekomgeving te diversifiëren, kunnen bacteriën over de hele planeet worden vervoerd, door wind of water, ze zijn erg licht en kunnen heel gemakkelijk worden meegenomen.
Indien nodig kunnen bacteriën afdichtingen vormen, door de ophoping van organismen, als ze een soort van obstakel willen organiseren of proberen opeengepakt te blijven. Microben worden op basis van enkele verschillen in groepen verdeeld.
Zo'n soort bacteriën als saprofyten voeden zich met de overblijfselen van dode dieren en planten, het is met hun hulp dat het vervalproces plaatsvindt.
Sommige bacteriën zijn alleen in staat om te groeien en zich te vermenigvuldigen onder de omstandigheden van een zuurstofomhulsel, zonder dat sterft het, en sommige soorten kunnen absoluut zonder zuurstof, de derde maakt helemaal geen verschil in welke habitat ze moeten bestaan, ze passen zich aan elke van hen.
Opgemerkt moet worden dat de anaërobe bacterie direct betrokken is bij de afbraak van eiwitten en vetten van dode planten en dieren. Het is dankzij hen dat het vervalproces plaatsvindt en een onaangename geur zich verspreidt.
Veel mensen vinden dit onaangenaam en moeilijk te verdragen, hoewel men moet bedenken dat het dankzij deze bacteriën is dat circulatie en uitwisseling in de natuur plaatsvinden, dode cellen splitsen en verdwijnen.
Zij waren het die een gunstige omgeving creëerden voor de verdere ontwikkeling van het leven, ze organiseerden de maliënkolders en turf, creëerden en handhaafden de circulatie van stoffen, ze balanceren ook de ideale verhouding van koolstofdioxide en zuurstof in de atmosfeer van de aarde.
Ongetwijfeld verergeren sommige microben ziekteprocessen en brengen ze meer infecties met zich mee, maar andere zijn betrokken bij het verteringsproces en het is niet bekend hoe het menselijk lichaam zou werken zonder de deelname van darmmicro-organismen.
De structuur en voeding van bacteriën

Bacteriën worden niet voor niets protozoa genoemd, hun eencellige structuur is echt heel laconiek. Het lichaam bestaat uit een binnenkern, waar alle biochemische processen en plasma plaatsvinden, die een beschermende functie vervult en schade weerstaat, en ook deelneemt aan het verkrijgen van voedingsstoffen, deze zelf doorgeeft, en verwerkte producten en gassen gemakkelijk naar buiten komen.
Onder bepaalde omstandigheden en habitat kan de cel zich ontwikkelen en zichzelf omringen met een speciale bescherming - plasma. Ze omringen de kooi niet altijd, maar alleen wanneer dat nodig is, zodat de kooi niet uitdroogt.
Het grootste deel van de cel is gevuld met een dikke vloeistof - dit is het cytoplasma.
Het is in lagen gerangschikt en alle voedingsstoffen vinden er hun plaats in. De kern is, zoals bij dierlijke cellulaire organismen, afwezig, maar de functies veranderen hierdoor niet. In het centrum van de cel is een stof geconcentreerd, begiftigd met erfelijke informatie en direct betrokken bij de reproductie van bacteriën.
Bacteriën voeden zich ook met totaal verschillende organismen, elke bacil is ontworpen om bepaalde stoffen te verwerken. Sommigen voeden zich met dode cellen en bevorderen hun verwerking, terwijl anderen juist levende cellen eten, wat grote schade toebrengt aan levende lichamen.
Microben die op planten leven, maken hun leven veel gemakkelijker door de wortels en het buitenste deel van de structuur te verwerken, wat grote voordelen met zich meebrengt, en bij het verwerken en gebruiken van koolstofdioxide wordt er een evenwicht bereikt in de atmosfeer van de planeet.
Beweging van bacteriën

Naast de chaotische beweging door middel van water en wind, hebben bacteriën zelf het recht om te kiezen waar ze zich heen willen bewegen.
Vraag: hoe verplaatsen bacteriën zich?
De locatie van de omringende flagella in bacteriën maakt het voor organismen mogelijk om zich langs het oppervlak te verplaatsen. Een massa bacteriën met tal van flagellen heeft het vermogen om zich in water en op het land te verplaatsen en bepaalt waar en met welk doel de bacteriën zullen gaan.
Meestal zijn bacteriële flagella dunne filamenten die beginnen bij het membraan van het bacteriële lichaam. Sommige flagellaten zijn uitgerust met een transportmiddel dat veel groter is dan het lichaam zelf, waardoor het mogelijk is om beter en sneller aan de oppervlakte te bewegen en door te stoten in een niet bijzonder gunstige omgeving of in een massa met een verzadigde bacteriële achtergrond.
Het kenmerk van beweging is vergelijkbaar met de rotatie van een propeller, dat wil zeggen, bij een bepaalde snelheid roteren de bacillen en dus vindt beweging plaats.
De locatie van flagella en hun aantal kunnen worden gekenmerkt door hun habitat: water of land. Micro-organismen die flagella over het hele oppervlak van het lichaam hebben, worden peritrichous genoemd.
Elke bacil heeft een andere lengte en dikte van het flagellum, maar van binnen is het altijd hol. Het volgende type monotrich, het voorvoegsel mono, spreekt voor zich en betekent dat het lichaam maar 1 flagellum heeft. Het is al veel moeilijker om ermee te bewegen.
Gecompliceerde micro-organismen hebben flagellen, in de vorm van microtubuli. Dit is een evolutionaire stap, en dergelijke bacteriën zijn een klasse hoger in hun lichaamsstructuur, en hun beweging is veel sneller en gemakkelijker.
De beweging van bacteriën vindt dus plaats door rotatie of duwen in een verdichte omgeving. Elke microbe is aangepast aan verschillende leefomstandigheden en habitats. Wetenschappers voerden veel analyses uit en kwamen tot de conclusie dat de beweging van microben intellectueel zinvol is en dat hun beweging in een opzettelijke richting plaatsvindt.
Het doel van microben in de natuur is veelzijdig, ze zijn veel eerder gecreëerd door de hele menselijke evolutie en hun aanwezigheid op de planeet is begiftigd met betekenis, sommige schaden levende organismen, maar toch bieden de meeste enorme hulp en ondersteuning in het bestaan van de planeet en de hele atmosfeer.
Flagella zijn lange draadvormige formaties op het oppervlak van de cel die zorgen voor de actieve ruimtelijke beweging. Ondanks de diversiteit aan organismen worden deze structuren binnen elk superrijk (prokaryoten of eukaryoten) gekenmerkt door een algemeen structuurschema.
Algemene kenmerken van flagella
In prenucleaire organismen (bacteriën en archaea) zijn flagellen de belangrijkste manier van bewegen. Onder eukaryoten zijn deze bewegingsstructuren voornamelijk aanwezig in eencellige organismen - protozoa, maar ze zijn ook kenmerkend voor gameten van planten en dieren. Bij sommige meercellige ongewervelde dieren, zoals sponzen, vervullen flagella de functie van het verplaatsen van een vloeibaar substraat ten opzichte van een onbeweeglijke cellaag.
Morfologisch bestaat het flagellum uit een basis die in de dikte van de cel is gefixeerd en een lange buitenste gloeidraad die roterende bewegingen langs een spiraalvormig traject maakt. De structuur en het werkingsmechanisme van deze delen in prokaryoten en eukaryoten zijn zeer verschillend, in verband waarmee twee overeenkomstige klassen van flagella worden onderscheiden.
Kenmerken van de flagella van prokaryoten en eukaryoten
Het buitenste filament van het flagellum wordt een filament genoemd. In prokaryoten bestaat het uit het flagelline-eiwit en beweegt het passief door de rotatie van de basale motor. Het filament van kerncellen is veel complexer en kan door de interactie van de eiwitten tubuline en dyneïne zelfstandig buigen.
| in prokaryoten | in eukaryoten | |
| organelgrootte (dikte, nm; lengte, m) | 10-30 nm, 6-15 micron | 200 nm, 100 m |
| filamenteuze eiwitten | flagelline | tubuline en dyneïne |
| membraan rond de gloeidraad | afwezig | is aanwezig |
| mate van rotatie | 360 ° | 180 ° |
| energiebron | transmembraanpotentiaal (in archaea, mogelijk ATP) | ATF |
| draad beweging | passief | actief |
| onderconstructies | filament, haak, basaal lichaam | filament, basaal lichaam (kinetosoom) |
| filamentstructuur: | vaste (in archaea) of holle eiwitcilinder | microtubuli doubletten |
| basale structuur | een staaf gefixeerd in een complex systeem van ringen of membraanachtige organellen (in archaea) | tripletten van microtubuli |
Een dergelijk aantal verschillen wijst op het gebrek aan homologie tussen deze organellen, dat wil zeggen dat ze niet hetzelfde zijn in oorsprong en structuur, hoewel ze vergelijkbare functies vervullen.
Het superrijk van prokaryoten omvat de koninkrijken van archaea en bacteriën. De locomotorische structuren van deze taxa zijn ook niet homoloog aan elkaar, maar lijken qua structuur sterk op elkaar. Archaea flagella zijn veel minder bestudeerd.
Archaea en bacteriën flagella
Volgens de bewegingsmethode zijn beweeglijke bacteriën verdeeld in zwevend en glijdend. Flagella zijn een bewegingsorgaan van drijvende micro-organismen, waardoor ze een snelheid van 20 tot 200 micron/sec kunnen ontwikkelen.
De beweging kan spontaan zijn (als de fysisch-chemische kenmerken van de omgeving in alle richtingen hetzelfde zijn) of doelgericht, wanneer de bacterie in de meest gunstige omstandigheden probeert te komen. Bij adaptieve beweging wordt de rotatie van de basale motor gestuurd door sensorische systemen.
Afhankelijk van het aantal en de locatie van flagella op de cel, onderscheiden bacteriën vier morfologische soorten micro-organismen:
- monotrichs - hebben een enkele flagellum;
- lofotrichi - gekenmerkt door een flagellaatbundel aan een van de celpolen;
- amphirichs - hebben een of meer flagellen aan beide uiteinden van de cel;
- peritrichous - bedekt met veel flagella aan alle kanten.

Het type geseling kan een soorteigenschap zijn of het gevolg zijn van een verandering in teeltomstandigheden of het stadium van de bacteriële levenscyclus.
Het archaeale flagellum lijkt in veel opzichten op het bacteriële flagellum, maar het heeft een aantal verschillen in zijn ultrastructuur en bewegingsmechanisme. Het filament in archaea is dus dunner, opgebouwd uit een ander type flagelline, en er is geen holle buis in het filament. De lengte van de haak is niet constant, het basale lichaam heeft een heel andere structuur en functies, hoogstwaarschijnlijk op basis van ATP-energie. Archaea bewegen veel langzamer dan bacteriën.
De structuur en functie van het bacteriële flagellum
Het bacteriële flagellum wordt gevormd door drie substructuren: het buitenste filament (filament), het flexibele gewricht (haak) en het basale lichaam, verankerd in het celmembraan. De synthese en assemblage van deze elementen wordt gecodeerd door ongeveer 50 fla-genen. De mot-genen zijn verantwoordelijk voor de werking van de motor en de che-genen zijn verantwoordelijk voor de adaptieve reacties.
Het filament van het flagellum is een relatief stijve eiwithelix die tegen de klok in is gedraaid om een centraal hol kanaal te vormen met een diameter tot 3 nm. Dit ontwerp draagt bij aan de vorming van een spiraalvormig traject van de draad. Flagelline-moleculen (FliC) worden langs het filamentkanaal getransporteerd.
De haak verbindt het filament met het basale lichaam van het flagellum en bestaat uit twee soorten eiwitten: FlgE en FlgKl. De articulatielengte is constant op ongeveer 50 nm. Door de gebogen vorm van de haak, wanneer de motor draait, maakt de basis van de fibril een cirkel, waardoor een spiraalvormige beweging van het flagellum mogelijk is.
Het basale lichaam is verankerd in de celwand en het cytoplasmatische membraan van bacteriën. Deze onderbouw heeft niet alleen een fixerende functie, maar is ook de motor van het flagellum. De structuur en lokalisatie van het basale lichaam hangen af van het type celwand van het micro-organisme. Bij gramnegatieve bacteriën bestaat het uit twee binnenste (M en S) en twee buitenste (P en L) ringen die aan een staaf zijn geregen die met een haak is verbonden. Het basale lichaam bevat ook een exportsysteem dat eiwitelementen transporteert voor de assemblage van het flagellum.
Basale motorstructuur
Het M-S-complex omvat bewegende structuren die een rotor worden genoemd en een draairichtingschakelaar, die in meer gedetailleerde bouwschema's een C-ring wordt genoemd. Rondom de rotor zijn door MotAB-eiwitten gevormde ionenkanalen - stators - geconcentreerd. De motor wordt aangedreven door de energie van een proton (H +) of natrium (Na +) gradiënt.

De rangschikking van cirkelvormige subeenheden in de celwand komt overeen met het volgende schema:
- "M" - cytoplasmatisch membraan;
- "S" - periplasmatische ruimte of celwand in G + -bacteriën;
- "P" - peptidoglycaanlaag;
- "L" is het buitenste membraan.
De buitenste ringen P en L zijn stationair en ondersteunend. Ze zijn afwezig in grampositieve bacteriën.
Nucleaire cel flagellum
Het eukaryote flagellum is een cytoplasmatische uitgroei van een cel, bestaande uit een buitenste deel omgeven door een membraan (undulipodium) en ondergedompeld in het cytoplasma van het basale lichaam (kinetosoom).

De structurele basis van undulipodium is het axoneme, dat bestaat uit een systeem van met elkaar verbonden eiwitcilinders - microtubuli. Hun opstelling komt overeen met de formule 9 × 2 + 2, dat wil zeggen negen perifere doubletten en twee enkele buizen in het midden (singlets).
De doubletten worden gevormd door gepaarde A- en B-cilinders die zijn opgebouwd uit tubuline-eiwitsubeenheden. Dynein-handgrepen strekken zich uit van elke A-buis naar het naburige paar, dat de energie van ATP omzet in mechanische beweging. De doubletten zijn verbonden met de singlets door radiale naalden en met elkaar door nexine ligamenten. De ruimte tussen de structurele elementen van undulipodium is gevuld met cytoplasma.

De structuur van het kinetosoom wordt weergegeven door negen tripletten van microtubuli (formule 9 + 0), die het flagellum verankeren in een eukaryote cel. Er zijn geen singlets in het basale lichaam.
Structuur. Ongeveer de helft van de bekende soorten bacteriën op het oppervlak heeft bewegingsorganen - golfachtige gebogen flagella. De massa van de flagella is tot 2% van de droge massa van de bacterie. De lengte van het flagellum is groter dan de lengte van het lichaam van het micro-organisme en is 3-12 µm; flagellum dikte 0,02 µm, met polaire flagella dikker dan peritrichous.
Flagella zijn samengesteld uit het eiwit flagelline (Latijnse flagella - flagellum), dat in zijn structuur verwijst naar contractiele eiwitten zoals myosine. Het flagellum bevat ofwel één homogeen eiwitfilament, of 2-3 filamenten die strak in een vlecht zijn gerold. Het filament van het flagellum is een stijve spiraal, tegen de klok in gedraaid; de toonhoogte van de spiraal is specifiek voor elk type bacterie.
Het aantal, de grootte en de locatie van flagella zijn kenmerken die constant zijn voor een bepaalde soort en waarmee rekening wordt gehouden in de taxonomie. Sommige bacteriën kunnen echter verschillende soorten flagella ontwikkelen. Bovendien hangt de aanwezigheid van flagella af van de omgevingsomstandigheden: op vaste media kunnen bacteriën bij langdurige teelt flagella verliezen en op vloeibare media kunnen ze deze opnieuw verwerven. Het aantal en de locatie van flagella bij dezelfde soort kan worden bepaald door het stadium van de levenscyclus. Daarom moet de taxonomische betekenis van deze eigenschap niet worden overschat.
Classificatie van bacteriën op basis van het aantal en de locatie van flagella:
1. Atrihs - flagellen ontbreken.
2. Monotrichs- een flagellum aan een van de polen van de cel (geslacht Vibrio)- monopolaire monotriche rangschikking van flagella, de meest mobiele bacteriën.
3. Politrichs - veel flagella:
– lophotrichs- een bundel flagella aan een pool van de cel (bevalling Pseudomonas, Burkholderia) - monopolaire polytricale rangschikking van flagella;
– amfitrix- aan elke pool van de cel bevindt zich een bundel flagellum (geslacht Spirillum)- bipolaire polytricale rangschikking van flagella;
– peritrichs- flagellen bevinden zich in willekeurige volgorde over het gehele oppervlak van de cel (fam. Enterobacteriën(bevalling Escherichia, Proteus), sem. Bacillaceae, dit Clostidiaceae), het aantal flagellen varieert van 6 tot 1000 per cel, afhankelijk van het type bacterie (Fig. 7).
Afb. 7. Varianten van de locatie van flagella in bacteriën:
1 - monotrich, 2 - lofotrich;
3 - amphitricus; 4 - peririchus.
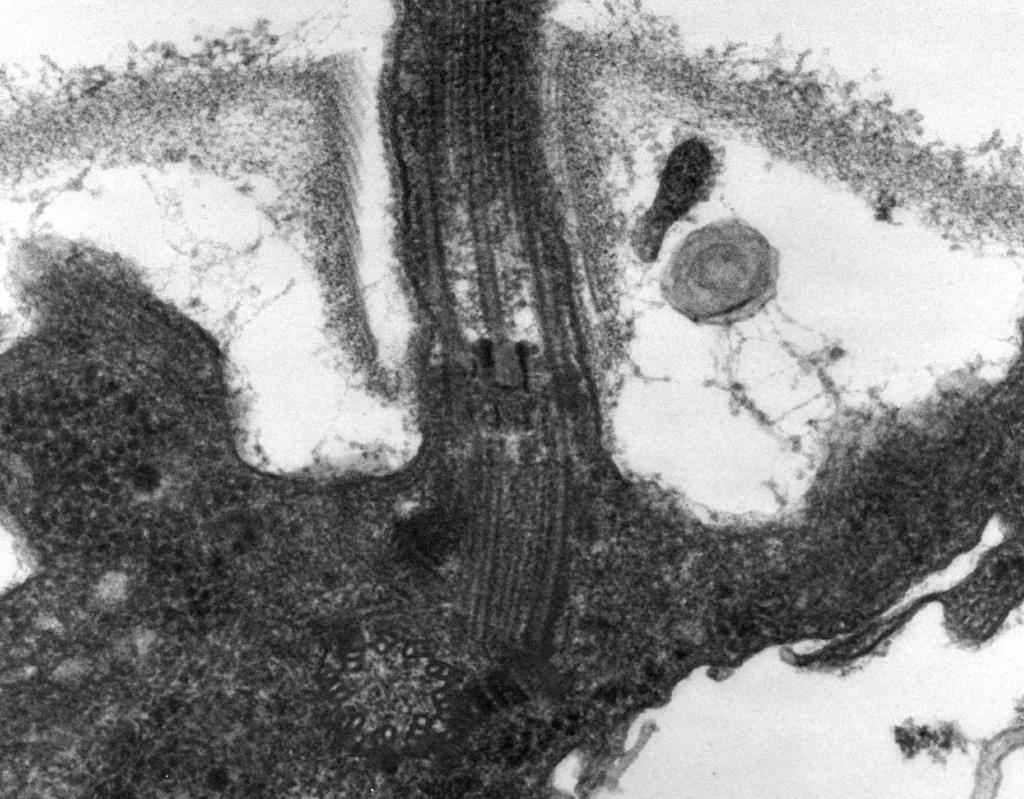
- Elektronenmicroscopie onthulde dat: het flagellum bestaat uit drie delen: spiraalfilament, haak en basislichaam (Fig. 8).
Het grootste deel van het flagellum is lang opgerolde draad (fibril) - een stijve holle cilinder met een diameter van ongeveer 120 nm, bestaande uit het flagelline-eiwit. Langs de lengte van de draad vormen eiwitmoleculen 11 rijen en zijn in een spiraal gerangschikt. Tijdens het proces van filamentgroei gaan eiwitmoleculen die in de cel worden gesynthetiseerd door de holte van de cilinder en worden aan het uiteinde aan de spiraal vastgemaakt. Aan het uiteinde van het flagellum bevindt zich een eiwitkap (dop) die de opening van de cilinder afsluit en voorkomt dat eiwitmoleculen in de omgeving vrijkomen. De lengte van het filament van het flagellum kan enkele micrometers bereiken. Bij sommige soorten bacteriën is het flagellum bovendien aan de buitenkant bedekt met een afdekking. Aan het oppervlak van de CS gaat de spiraalvormige draad over in een verdikte gebogen structuur - een haak.

Rijst. acht. Diagram van de structuur van het flagellum
2. Haak(20-45 nm dik) nabij het celoppervlak - een relatief korte cilinder, bestaat uit een ander eiwit dan flagelline en dient voor een flexibele verbinding van het filament met het basale lichaam.
3. Basaal lichaam bevindt zich aan de basis van het flagellum en zorgt voor de rotatie ervan. Het basale lichaam bevat 9-12 verschillende eiwitten en bestaat uit twee of vier schijven (ringen) die aan een staaf zijn geregen, die een voortzetting is van de haak. Deze ringen zijn ingebouwd in de CPM en KS. De twee binnenringen (M en S) zijn essentiële componenten van het basale lichaam. De M-ring bevindt zich in de CPM, de S-ring bevindt zich in de periplasmatische ruimte van gramnegatieve bacteriën of in de peptidoglycaanzak van grampositieve bacteriën. Twee buitenste ringen (D en L) zijn optioneel voor beweging, omdat ze alleen aanwezig zijn in gramnegatieve bacteriën, respectievelijk gelokaliseerd in de peptidoglycaanlaag en in het buitenmembraan van de CS. Ringen S, D en L zijn onbeweeglijk en dienen om het flagellum in het kniegewricht te fixeren. De rotatie van de flagellum wordt bepaald door de rotatie van de M-ring die in de cel CPM is ingebouwd. De structurele kenmerken van het basale lichaam van het flagellum worden dus bepaald door de structuur van de CS.
Functioneel gezien is het basale lichaam een elektromotor die wordt aangedreven door protonen. M-ring van het basale lichaam (roterende rotor), omgeven door membraaneiwitten met negatieve ladingen (motorstator). De bacteriële cel heeft een effectief mechanisme voor het omzetten van elektrochemische energie in mechanische energie. Daarom besteedt de bacterie ongeveer 0,1% van alle energie die hij verbruikt aan het werk van het flagellum. Tijdens de werking van het flagellum wordt een proton-aandrijvende kracht gebruikt, die wordt geleverd door het verschil in de concentratie van protonen aan de buiten- en binnenkant van het membraan (er zijn er meer aan de buitenkant) en de aanwezigheid van een meer negatieve lading aan de binnenkant van het membraan. De proton-aandrijvende kracht dwingt protonen om door het basale lichaam de cel in te gaan, terwijl ze in bepaalde delen van de rotor blijven, waardoor ze een positieve lading krijgen, en dan gaan de protonen de cel in. De geladen secties zijn zo geplaatst dat er een aantrekkingskracht ontstaat tussen de geladen secties van de rotor en stator, de M-ring begint te draaien met een snelheid van ongeveer 300 toeren/sec. Rotatiemechanisme: opladen - opladen van de COOH-groep in aminozuren. Voor een volledige rotatie van de ring moeten 500-1000 protonen door het basale lichaam gaan. De rotatie van de M-ring door een star verbonden as en de haak wordt overgebracht op het filament van het flagellum, dat fungeert als propeller of scheepsschroef. De bacterie drijft zolang de propeller draait, de bijdrage van traagheid is extreem klein.
Bovendien bewegen bacteriën, zelfs dode, die zich in het aquatisch milieu bevinden, als gevolg van Brownse beweging. De bacteriecel is voortdurend onderhevig aan invloeden van omringende moleculen in thermische beweging. Inslagen uit verschillende richtingen gooien de bacteriën heen en weer.
Het type beweging van de flagella is rotatie. Er zijn twee soorten bewegingen: rechtlijnig en salto (periodieke willekeurige veranderingen in bewegingsrichting). Wanneer de filamenten tegen de klok in draaien (ongeveer 1 seconde), met een frequentie van 40-60 rpm / s (dicht bij de snelheid van een gemiddelde elektromotor), worden hun draden tot een enkele bundel geweven (figuur 9a). De rotatie van de flagella wordt doorgegeven aan de cel. Omdat de cel veel massiever is dan het flagellum, begint deze in een rechte lijn in de tegenovergestelde richting te bewegen, met een snelheid die 3 keer lager is dan de snelheid van het flagellum.
Dit zorgt voor de translatiebeweging van de cel, waarvan de snelheid in een vloeibaar medium voor verschillende soorten bacteriën 20-200 µm/s is (dit komt overeen met ongeveer 300-3000 lichaamslengtes per minuut) en een langzamere beweging over het oppervlak van vaste stof. media.
Een bacterie kan doelbewust maximaal 3 seconden in één richting zwemmen, waarna de inslagen van omringende moleculen hem in een willekeurige richting draaien. Daarom hebben bacteriën een mechanisme ontwikkeld voor spontane verandering in bewegingsrichting - het schakelen van de flagellaire motor. Wanneer het met de klok mee begint te draaien (ongeveer 0,1 s), stopt de bacterie en draait zich om (maakt een "rol") in een willekeurige richting. In dit geval vliegen de flagellen in verschillende richtingen uit elkaar (Fig. 9b). Bij amfitroenen wordt tijdens het bewegen een bundel flagella binnenstebuiten gekeerd (zoals een paraplu die door de wind wordt gedraaid). Dan schakelt de motor weer tegen de klok in en drijft de bacterie weer in een rechte lijn, maar in een andere, willekeurige richting.
Flagella kan ook van richting veranderen als reactie op externe prikkels. Als de bacterie naar de optimale concentratie van de lokstof beweegt, duwen de flagellen de cel door het medium, de rechtlijnige beweging wordt langer en de tuimelfrequentie is lager, waardoor deze uiteindelijk in de gewenste richting kan bewegen.
Er zijn gevallen bekend van het bestaan van inactieve (verlamde) flagella. Voor de beweging van flagellaire bacteriën is intactheid (intactheid) van de CS vereist. De behandeling van cellen met lysozyme, dat de peptidoglycaanlaag van het CS verwijdert, zorgt ervoor dat de bacteriën hun vermogen om te bewegen verliezen, hoewel de flagellen intact blijven.
Taxi's van bacteriën. Zolang de omgeving onveranderd blijft, drijven bacteriën willekeurig rond. Een volledig homogene omgeving is echter zelden het geval. Als de omgeving heterogeen is, vertonen bacteriën elementaire gedragsreacties: ze bewegen actief in de richting die bepaald wordt door bepaalde externe factoren. Dergelijke genetisch bepaalde gerichte bewegingen van bacteriën worden taxi's genoemd. Afhankelijk van de factor onderscheidt men chemotaxis (een speciaal geval is aerotaxis), fototaxis, magnetotaxis, thermotaxis en viscositaxis.
chemotaxis- beweging in een specifieke richting ten opzichte van de bron van de chemische stof. Chemicaliën zijn onderverdeeld in twee groepen: inerte en taxiveroorzakende - chemo-effectoren. Onder chemo-effectors zijn er stoffen die bacteriën aantrekken - lokstoffen (suikers, aminozuren, vitamines, nucleotiden) en stoffen die ze afstoten - insectenwerende middelen (sommige aminozuren, alcoholen, fenolen, anorganische ionen). Een lokstof voor aërobe en een afweermiddel voor anaërobe prokaryoten is moleculaire zuurstof. Lokmiddelen worden vaak gepresenteerd als voedingssubstraten, hoewel niet alle voor het lichaam noodzakelijke stoffen als lokstoffen werken. Ook dienen niet alle giftige stoffen als insectenwerende middelen en zijn niet alle insectenwerende middelen schadelijk. Bacteriën kunnen dus niet op verbindingen reageren, maar alleen op bepaalde en voor verschillende bacteriën verschillend.
In de oppervlaktestructuren van een bacteriële cel bevinden zich speciale eiwitmoleculen - receptoren die specifiek binden aan een bepaalde chemo-effector, terwijl het chemo-effectormolecuul niet verandert, maar conformationele veranderingen optreden in het receptormolecuul. Receptoren bevinden zich ongelijkmatig over het gehele oppervlak van de cel en zijn geconcentreerd op een van de polen. De toestand van de receptor weerspiegelt de extracellulaire concentratie van de overeenkomstige effector.
Chemotaxis is adaptief. Vormen van Vibrio cholerae met verminderde chemotaxis zijn bijvoorbeeld minder virulent.
Aerotaxis- Bacteriën die moleculaire zuurstof nodig hebben, hopen zich op rond luchtbellen die onder het dekglas zijn opgesloten.
Fototaxi- beweging naar of weg van licht is kenmerkend voor fototrofe bacteriën die licht gebruiken als energiebron.
Magnetotaxis- het vermogen van waterbacteriën die kristallen van ijzerbevattende mineralen bevatten om langs de lijnen van het aardmagnetisch veld te zwemmen.
Thermotaxis- beweging in de richting van temperatuurverandering, wat van groot belang is voor sommige pathogene bacteriën.
Viscositaxis- het vermogen om te reageren op veranderingen in de viscositeit van de oplossing. Meestal neigen bacteriën naar een medium met een hogere viscositeit, wat van groot belang is voor pathogene soorten.
Slip bacteriën. Het vermogen om met een lage snelheid (2-11 m / s) op een vast of stroperig substraat te glijden, is gevonden in sommige prokaryoten, bijvoorbeeld mycoplasma's.
Er zijn verschillende hypothesen om de glijdende beweging te verklaren. Volgens straalvoortstuwingshypothese het wordt veroorzaakt door de afscheiding van slijm door talrijke slijmporiën in het CS, waardoor de cel wordt afgestoten van het substraat in de richting tegengesteld aan de richting van de slijmafscheiding. Volgens lopende golf hypothese glijdende beweging in mobiele flagella-loze vormen wordt geassocieerd met de aanwezigheid tussen de peptidoglycaanlaag en het buitenmembraan van de CS van een dunne eiwitlaag van geordende fibrillen, vergelijkbaar met de filamenten van flagella. De roterende beweging van de fibrillen, "getriggerd" door deze structuren, leidt tot het verschijnen van een "reizende golf" (bewegende microscopisch kleine uitstulpingen van de CS) op het celoppervlak, waardoor de cel wordt afgestoten van het substraat. Ten slotte zijn in sommige glijdende bacteriën structuren beschreven die lijken op de basale lichamen van flagellaire vormen.
Functies van flagella:
1. Zorg voor hechting - de beginfase van het infectieuze proces.
2. Zorg voor de mobiliteit van bacteriën.
3. Bepaal de antigene specificiteit, het is het H-antigeen.
Identificatie van flagella:
1. Fasecontrastmicroscopie van inheemse preparaten ("gemalen" en "hangende" druppels). Microscopisch wordt de beweeglijkheid bepaald in cellen van dagelijkse kweek. Om mobiliteit te onderscheiden van passieve Brownse beweging, wordt een druppel van een 5% waterige oplossing van fenol toegevoegd aan een druppel van de onderzochte cultuur; actieve beweging stopt in dit geval.
2. Donkerveldmicroscopie van inheemse preparaten.
3. Lichtmicroscopie van met kleurstoffen of metalen bevlekte preparaten. Omdat flagella tijdens de bereiding heel gemakkelijk beschadigd raakt, worden deze methoden in de dagelijkse praktijk zelden gebruikt.
Voor het kleuren van flagella worden op schuine agar gekweekte cellen gebruikt. Met een bacteriële lus worden cellen uit het condenswater gehaald en voorzichtig overgebracht in steriel gedestilleerd water van dezelfde temperatuur als de incubatietemperatuur van bacteriën op de agarhelling, en de bacteriën worden niet van de lus geschud, maar zorgvuldig ondergedompeld in water. Het reageerbuisje met bacteriën wordt 30 minuten op kamertemperatuur gelaten. Gebruik chemisch zuiver (gewassen in een chroommengsel) glas, waarop 2-3 druppels suspensie worden aangebracht. De suspensie wordt verspreid over het glasoppervlak door het voorzichtig te kantelen. Droog het preparaat aan de lucht.
Flagella zijn erg dun, dus ze kunnen alleen worden gedetecteerd met een speciale verwerking. Eerst wordt met behulp van etsen zwelling en vergroting van hun grootte bereikt, en vervolgens wordt het preparaat gekleurd, waardoor ze zichtbaar worden onder lichtmicroscopie.
Gebruik vaker verzilveringsmethode volgens Morozov (fig. 10):
- het preparaat wordt gedurende 1 minuut gefixeerd met een oplossing van ijsazijn, gewassen met water;
- breng een oplossing van tannine aan (bruinen, maakt de flagella dichter) gedurende 1 minuut, gewassen met water;
- het preparaat wordt behandeld door 1-2 min te verwarmen met een impregneeroplossing van zilvernitraat, gewassen met water, gedroogd en gemicroscoop.
Microscopie toont donkerbruine cellen en lichtere flagellen.

Rijst. tien. Identificatie van flagella door de methode van verzilvering
Rijst. elf. Identificatie van flagella
door elektronenmicroscopie
4. Elektronenmicroscopie van met zware metalen besproeide preparaten (Fig. 11).
5. Indirect - door de aard van de groei van bacteriën wanneer geïnoculeerd in halfvloeibare 0,3% agar. Na incubatie van gewassen in een thermostaat gedurende 1-2 dagen, wordt de aard van bacteriegroei opgemerkt:
- in immobiele bacteriën (bijvoorbeeld S. saprophyticus) er is een toename in de loop van de injectie - "nagel", en het medium is transparant;
- in beweeglijke bacteriën (bijv. E coli) er is groei aan de zijkanten van de injectie, langs de hele kolom van agar - "visgraat", en diffuse troebelheid van het medium.